Computational and Systems Neuroscience (COSYNE) II-37, 2013.
Urs Bergmann, Michiel Remme, Susanne Schreiber, Henning Sprekeler and
Richard Kempter
Declarative memories initially depend on the hippocampus. Over a period of weeks to years, however, these memories become hippocampus-independent through a process called system memory consolidation. The underlying cellular mechanisms are unclear. Here, we suggest a consolidation mechanism based on an anatomical network motif and spike timing-dependent plasticity. As a first step in the memory consolidation process, we focus on pyramidal neurons in the hippocampal CA1 area. Information from entorhinal cortex reaches these cells through two pathways: an indirect pathway via CA3 and the Schaffer collaterals (SC), and the direct perforant path (PP). Memory patterns are assumed to be initially stored in the recurrent CA3 network and SC synapses during the awake exploratory state. During a subsequent consolidation phase, CA3/SC-dependent memory patterns are then partly transfered to the PP synapses. Using numerical simulations and mathematical analysis, we show that this consolidation process occurs as a natural result from the combination of (1) spike timing-dependent plasticity at PP synapses and (2) the temporal correlations between SC and PP activity, since the SC input is delayed compared to the PP input (5-15 ms). Investigation of alternating wake and sleep phases reveals a decay of the memories in SC, but a slower decay in the PP connection to CA1. Less memory consolidation to the PP allows longer memory retention times, but comes at the price of worse initial memory storage. In a hierarchical network model that repeats the network motif across many levels, each direct connection at one level is part of the indirect pathway of the next level. Learning rates decrease with increasing level. Theoretical analysis and simulations of the hierarchical system show power-law forgetting, as seen with psychophysical forgetting functions (Wixted and Ebbesen 1991). Furthermore, consolidated memories yield faster responses because they are stored in increasingly shorter synaptic pathways.
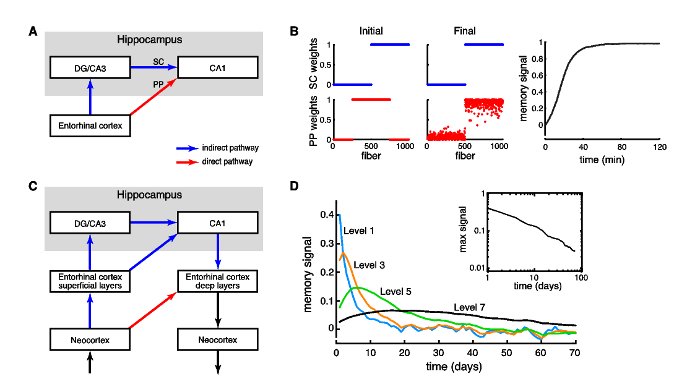
Figure 1: A: Schematic of hippocampal network with entorhinal cortex projecting directly to CA1 cells via the perforant path (PP; red arrow), and indirectly (blue arrows) via the dentate gyrus and CA3 through the Schaffer collaterals (SC). B: Simulation with 1000 SC fibers and 1000 PP fibers connecting to a single CA1 cell (leaky integrate-and-fire). The PP fibers show spike timingdependent plasticity (as in Song, Miller and Abbott, 2000). Left column shows the initial synaptic weights of the SC fibers (top) and PP fibers (bottom). Middle column shows the final weight distributions. Right panel shows the evolution of the memory signal in the PP (i.e. correlation between the SC and PP weight vector). C: At each level in the hierarchical model, the indirect pathway includes the lower level direct pathway(s). This indirect(blue)-direct(red) network motif is iterated over several levels. D: Memory signal decay is shown for increasingly slower learning rates for higher levels in a network of leaky integrate-and-fire neurons. A traveling memory wave can be observed. Inset: the maximum memory signal yields a power-law forgetting function.
Acknowledgments
Supported by BMBF grants no. 01GQ0901, 01GQ0972, 01GQ1201, 01GQ1001A, DFG grant no.
SFB 618 and the Einstein Stiftung Berlin.
Download Poster: JPG
{kind=link}